Temperature control of vesicular trafficking
Cells interact with their environment by secreting or importing proteins. These proteins are packaged in vesicles going in and out of the cell. Inside the cells, perfect trafficking organization ensures that vesicles are directed to the right place. The development of microscopy techniques, now allow biologists to track vesicles and decipher the role of motor proteins in vesicular transport using real-time imaging.
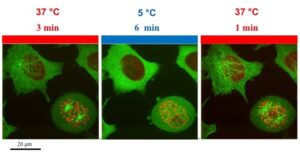
Our temperature controller allows you to precisely control the temperature of your cells, or switch temperature from 5 to 45C in seconds and thermally control vesicles entry, exit and transport inside the cells, take a tour!
Temperature control of vesicular trafficking
Vesicular trafficking is a process spanning from the cellular membrane to the cell organelles. At the membrane, endocytosis and exocytosis ensure cell communication with the environment, it also ensures receptors, ions channels, surface proteins and membrane lipids renewal. In the cytoplasm, vesicles travel along microtubules using motor proteins to shuttle to recycling organelles.
Endocytosis
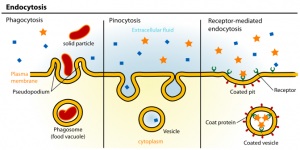
Endocytosis allows the cell to sense its environment by internalizing nutrients, signaling molecules and their receptors, or fight against bacteria. There are several mechanisms of endocytosis (see figure), one of the most detailed pathway is clathrin-mediated endocytosis. When a ligand binds to its surface receptor, a signal is sent inside the cell to recruit clathrin proteins to the membrane area to be endocytosed. Clathrin is composed of two subunits: a 190 kDa clathrin heavy chain and a 25 kDa light chain. Clathrin proteins arrange in a characteristic triskelion composed of three clathrin heavy chains and three clathrin light chains. Clathrin triskelions bind adaptor proteins on the forming vesicle and act by mechanically deforming the membrane to form a rounded vesicle. Subsequently, the membrane invaginates to form a small-coated pit, which will further grow into a coated vesicle. Vesicle/membrane separation is ensured by dynamin, a GTPase protein, forming an helix around the neck of the vesicle to pinch off the coated vesicle from the membrane. Once the vesicle is separated from the membrane, clathrin proteins disassemble from the vesicle. The vesicle is then shuttle to and fuse with a structure of the endocytic pathways called the early endosome. Early endosome have low pH lumen, which enable the dissociation of the endocytosed ligand/receptors complex and the recycling of the receptors.
Ultra fast temperature shift device for in vitro experiments under microscopy
Exocytosis
Exocytosis is a process allowing secretion of proteins, hormones, neurotransmitters, turnover of plasma membrane glycoproteins and receptors, and antigen presentation during the immune system reaction.
There are two mediated pathways for exocytosis: calcium-mediated, notably in neurons for neurotransmitters release, and a non-calcium mediated pathways, which is commonly used by cells to release exocytic vesicles.
Exocytosis is a four steps process: approach to plasma membrane, vesicle tethering, vesicle docking and vesicle fusion. Vesicles approach the plasma membrane by using motor proteins, such as kinesin or dynein, advancing on a microtubule track. When the vesicle is near the membrane it will loosely attach via the binding of Rab-GTP on the vesicle surface to Rab effector protein at the target membrane surface. Vesicles docking and fusion to the membrane is ensured by SNARE proteins. There are mainly two types of SNARE proteins: vesicle v-SNARE: synaptobrevin and target t-SNARE: syntaxin, SNAP25. The binding of v-SNARE with t-SNARE triggers the fusion of the membrane vesicles to the plasma membrane.
Motor protein
In the cytoplasm, vesicles are actively transported by motor proteins, which use microtubules as intracellular tracks. Motor proteins are molecular motors, which provide a mechanical force, via ATP hydrolysis, to ensure vesicle transport. There are two main types of microtubules motor proteins kinesin and dynein. Kinesin role was very well studied in neurons, where it ensures the transport of synaptic vesicles along the axon microtubules. Kinesin is composed of two globular head domains in contact with microtubules, a coiled-coil alfa helix and a light chain domain in contact with the vesicles. Kinesin proteins move from the (-) end towards the (+) ends of microtubules, while dynein transport vesicles from the (+) end towards the (-) ends, this direction of transport is needed for the retrograde transport of synaptic vesicles in neurons.

Temperature control of vesicular trafficking
Endocytosis, exocytosis or vesicular transports are very dynamic. Coated pits have a 1 min lifetime before they elongate into vesicles, and coated vesicles have an even shorter lifetime that can be measure in seconds (Kirchhausen, 2009). It is clear, that the available tools to study these processes have to be as dynamic as the processes themselves. With the development of TIRF microscopy and a wide-range of fluorescence proteins, it is now possible to track single vesicle in a cell. Vesicular trafficking is highly sensitive to temperature. Vesicular transport is slowed-down at temperature below 19C and is completely blocked at 4C.
Our temperature controller can switch temperature from 5 to 45C in seconds and is very precise. It offers the possibility to combine thermo-control of vesicles trafficking with high-resolution imaging.
References
Alberts B, Johnson A, Lewis J, Raff M, Roberts K, and Walter P. Molecular biology of the cells, New York: ncbi; 2002
Kirchhausen T, Imaging endocytic clathrin structures in living cells, Trends in Cell Biol, 2009
Hong G, Wu JZ, Robinson JT, Wang H, Zhang B and Dai HThree-dimensional imaging of single nanotube molecule endocytosis on plasmonic substrates, 2012
Cai D, McEwen D, Martens J, Meyhofer E, Verhey K. “Single Molecule Imaging Reveals Differences in Microtubule Track Selection Between Kinesin Motors”. PLOS Biology, 2009.
See also :
Cell culture and cell lines : https://en.wikipedia.org/wiki/Cell_culture
FAQ
Vesicular trafficking is a process that spans from the cellular membrane to the cell’s organelles. Cells interact with their surroundings by secreting or importing proteins. These proteins are packaged in vesicles. At the membrane, endocytosis and exocytosis permit communication with the environment. These actions also ensure the renewal of receptors, ion channels, surface proteins, and membrane lipids. Inside the cell, an organized trafficking system ensures vesicles are directed to the correct place. In the cytoplasm, vesicles travel along microtubules. They use motor proteins to shuttle to different organelles.
This is one of the most detailed pathways for endocytosis, which allows the cell to internalize substances. The process begins when a ligand binds to its receptor on the cell surface. A signal is then sent inside the cell to recruit clathrin proteins. Clathrin proteins are arranged in a triskelion shape. These triskelions bind to adaptor proteins on the forming vesicle. They function by mechanically deforming the membrane. This action forms a rounded vesicle. The membrane invaginates into a small coated pit, which then grows into a coated vesicle. A GTPase protein called dynamin ensures the vesicle separates from the membrane. Dynamin forms a helix around the vesicle’s neck to pinch it off. After separation, the clathrin proteins disassemble from the vesicle. The vesicle is then shuttled to the early endosome.
Exocytosis is a process that allows for the secretion of proteins, hormones, and neurotransmitters. It is also involved in antigen presentation and the turnover of plasma membrane components. The process involves four steps: approach, tethering, docking, and fusion. First, vesicles approach the plasma membrane, using motor proteins like kinesin or dynein that move on a microtubule track. Second, the vesicle loosely attaches to the membrane in a step called tethering. This occurs when Rab-GTP on the vesicle binds to a Rab effector protein at the target membrane. Third and fourth, vesicle docking and fusion are managed by SNARE proteins. There is a v-SNARE (vesicle SNARE) and t-SNAREs (target SNAREs). The binding of the v-SNARE to the t-SNAREs causes the fusion of the vesicle to the plasma membrane.
Vesicles are actively transported within the cytoplasm by motor proteins. These proteins are described as molecular motors. They use microtubules as intracellular tracks to move along. Motor proteins provide a mechanical force to ensure vesicle transport, a process that is fueled by ATP hydrolysis. The two main types of these proteins are kinesin and dynein. Kinesin proteins move from the minus end of the microtubules toward the plus end. This protein’s function was studied in neurons, where it transports synaptic vesicles along the axon. Dynein transports vesicles in the opposite direction, from the plus end toward the minus end. This direction of movement is necessary for retrograde transport in neurons.