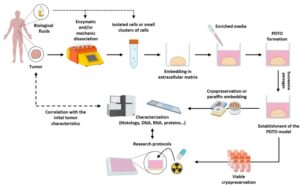
Adipocyte Interactions and Immunomodulation in Triple-Negative Breast Cancer
Adipocyte Transformation in the Tumour Microenvironment
Triple-negative breast cancer (TNBC) is defined by a lack of estrogen receptor, progesterone receptor, and human epidermal growth factor receptor 2 (HER2) expression [1]. This subtype is known for high metastatic potential and poor prognosis compared to other forms of breast cancer [1]. Within the tumour microenvironment (TME), which is the ecosystem comprising cancer cells, immune cells, and stromal components, adipocytes are a major constituent [2]. Adipocytes serve as energy storage units and endocrine cells [2]. In the presence of cancer cells, these adipocytes are transformed into cancer-associated adipocytes (CAA) [1].
CAA are observed at the invasive front of human breast tumours [2]. These cells exhibit distinct morphological changes, such as a smaller size and irregular shapes [2]. Lipid accumulation is reduced in CAA compared to normal mature adipocytes [1]. In laboratory models, CAA are established by co-culturing 3T3-L1-induced adipocytes with 4T1 breast cancer cells [1]. These cells lose markers of mature adipocyte differentiation, such as peroxisome proliferator-activated receptor gamma (PPAR-γ), CCAAT enhancer-binding protein alpha (C/EBPα), and fatty acid binding protein 4 (FABP4) [2, 7]. Simultaneously, markers of undifferentiated cells, such as alpha-smooth muscle actin (α-SMA) and vimentin, are increased [2, 7].
Adipose-derived mesenchymal stem cells (ADMSC) also respond to the TNBC secretome [6]. These cells can differentiate into several lineages, but when exposed to media from MDA-MB-231 cells, they acquire a CAA-like phenotype [6]. This transition involves the acquisition of a pro-inflammatory state characterised by the secretion of many cytokines [6]. The presence of crown-like structures (CLSs), which consist of macrophages surrounding dying adipocytes, is a histologic marker of this pro-inflammatory transition [2].
Signalling Pathways and Epithelial-Mesenchymal Transition
The progression of TNBC is often driven by the epithelial-mesenchymal transition (EMT), a sequence where epithelial cells lose polarity and acquire mesenchymal traits [1]. CAA promote this transition through the activation of several intracellular pathways [1]. One such pathway is the PI3K/AKT signalling cascade [1]. In tumour tissues where CAA are present, E-cadherin expression is downregulated, while Ki67 expression is increased, indicating higher proliferation [1].
Another major pathway involved in CAA-mediated progression is the Stat3 (signal transducer and activator of transcription 3) pathway [7]. Granulocyte colony-stimulating factor (G-CSF), secreted by CAA, activates Stat3 in TNBC cells [7]. This activation leads to the expression of pro-invasive genes, such as matrix metalloproteinase 2 (MMP2) and matrix metalloproteinase 9 (MMP9) [7]. Laboratory data display that targeting the G-CSF/Stat3 axis with neutralizing antibodies or siRNAs can abrogate the migration and invasion induced by CAA [7].
Snail is a biomarker that controls the acquisition of the CAA phenotype in ADMSC [6]. When ADMSC are exposed to the TNBC secretome, Snail expression is induced [6]. This induction is associated with the activation of Smad2 and NF-κB pathways [6]. These pathways coordinate the expression of inflammatory and immunomodulatory genes that support a tumour-promoting environment [6]. The use of epigallocatechin-3-gallate (EGCG), a polyphenol from green tea, has been shown to inhibit the activation of these pathways and prevent the onset of the CAA phenotype [6].
Cytokine-Mediated Communication and Metabolic Changes
CAA act as endocrine and paracrine cells by releasing a variety of cytokines and chemokines, collectively called adipokines [7]. These substances comprise interleukin-6 (IL-6), interleukin-8 (IL-8, also known as CXCL8), interleukin-1 beta (IL-1β), and C-C motif chemokine ligand 5 (CCL5) [2]. IL-6 is a pleiotropic cytokine that stimulates the invasion and migration of breast tumour cells through the Cofilin-1 pathway [2]. Other members of the IL-6 family, such as leukaemia inhibitory factor (LIF) and oncostatin M (OSM), also participate in metastatic transition and the degradation of the extracellular matrix (ECM) [2].
CXCL8 is a chemokine secreted by CAA that has been linked to tumour growth and metastasis [3]. High levels of CXCL8 are found in the secretome of CAA isolated from TNBC patients [3]. This chemokine promotes the proliferation and migration of cancer cells [3]. Separately, CCL2 and CCL5 are induced in ADMSC by the TNBC secretome [6]. These chemokines are associated with the recruitment of immune cells and the promotion of an invasive phenotype in cancer cells [2, 6].
Metabolic changes are also a feature of the interaction between cancer cells and adipocytes [2]. CAA provide metabolic substrates, such as free fatty acids (FFAs), to fuel the growth of cancer cells [1, 2]. This metabolic rewiring allows cancer cells to survive in the hypoxic conditions often found within solid tumours [2]. The secretion of metabolites like lactate, pyruvate, and ketone bodies by CAA further supports this adaptive state [6]. These changes contribute to an environment that facilitates both tumour expansion and the evasion of therapeutic agents [2].
Modification of the Tumour Immune Microenvironment
The tumour immune microenvironment (TIME) is a complex system of immune cells that can either inhibit or promote tumour growth [3]. CAA contribute to an immunosuppressive TIME by modulating the function of T cells and other immune populations [1]. One mechanism is the upregulation of programmed death-ligand 1 (PD-L1, also known as CD274) on the surface of TNBC cells [1, 3]. Increased PD-L1 expression allows cancer cells to evade detection by the immune system [1].
In tumours co-existing with CAA, a reduction in the infiltration of CD8+ T cells is observed [1]. These cells are the primary effectors of the anti-tumour immune response. The secretome of CAA, particularly CXCL8, has been shown to suppress both CD4+ T and CD8+ T cell infiltration [3]. For this reason, targeting the CXCL8 pathway may be a strategy to enhance the efficacy of immune checkpoint inhibitors, such as anti-PD-1 therapies [3]. In mice, the combination of a CXCL8 inhibitor and anti-PD-1 therapy resulted in a synergistic inhibition of tumour progression [3].
Immune evasion is also supported by the recruitment of immunosuppressive cells [2]. CAA-derived cytokines stimulate the recruitment of macrophages and promote their polarisation into a pro-tumour M2 phenotype [2]. This recruitment is mediated by factors such as CCL2 and VEGF [2]. The resulting environment is one where anti-tumour immunity is suppressed, and pro-inflammatory signals continue to drive malignancy [1, 2].
Extracellular Vesicles and Paracrine Regulation
Extracellular vesicles (EVs) are small, lipid-delimited particles that facilitate cell-to-cell communication by transferring proteins, lipids, and nucleic acids [4]. Exosomes are a subtype of EVs with a diameter between 30 and 150 nm [4]. TNBC cells, such as the MDA-MB-231 line, secrete EVs that can pre-condition the surrounding adipose tissue [4]. These vesicles trigger a pro-inflammatory phenotype in ADMSC, characterised by increased expression of CXCL8, CCL2, and IL-1β [4].
Recent data indicate that EVs from TNBC cells also carry mitochondrial components and mitochondrial DNA (mtDNA) [4]. The horizontal transfer of these components can modulate the metabolic function and viability of recipient cells [4]. In ADMSC, exposure to TNBC-derived EVs induces markers of senescence, such as p21 and beta-galactosidase [4]. This senescence is associated with the activation of signaling pathways like AKT and GSK-3β [4].
Dietary polyphenols like EGCG can alter the content and effect of these vesicles [4]. Treatment of MDA-MB-231 cells with EGCG modifies the genetic material found within their secreted EVs [4]. EGCG-treated EVs have a lower mitochondrial content and a reduced capacity to induce inflammation and senescence in ADMSC [4]. This suggests that diet-derived substances can interfere with the paracrine regulation exerted by TNBC cells on their local environment [4].
Single-Cell Analysis of Adipocyte Heterogeneity
White adipose tissue (WAT) is a metabolic organ found at several anatomic sites, including subcutaneous adipose tissue (SAT) and visceral adipose tissue (VAT) [5]. It comprises mature adipocytes and adipose stem and progenitor cells (ASPCs) [5]. Single-nucleus RNA sequencing (snRNA-seq) and single-cell RNA sequencing (scRNA-seq) have allowed for a detailed mapping of these cell populations in both mice and humans [5]. These methods reveal that adipocytes in the tumour-adipose microenvironment (TAME) are highly heterogeneous [5].
Several subtypes of cancer-associated adipocytes have been identified [5]. DPP4+ adipocytes are found in VAT, while ADIPOQ+ adipocytes are present in SAT [5]. High levels of these subtypes are associated with poor outcomes in adipose-associated cancers, including breast cancer [5]. These subtypes interact with immune cells and other stromal components to create an ecosystem that supports tumour development [5].
Bioinformatics screening of single-cell data has identified possible drugs to target these adipocyte subtypes [5]. These comprise Trametinib, Selumetinib, and Ulixertinib [5]. Separately, the knockdown of adiponectin receptor 1 (AdipoR1) and AdipoR2 has been shown to impair the proliferation and invasion of breast cancer cells in vitro [5]. Patients with high expression of AdipoR2 in breast cancer tissues have been observed to have shorter relapse-free survival compared to those with low expression [5]. This knowledge provides a basis for the development of targeted therapies that focus on exact adipocyte subpopulations within the TME [5].
Experimental Models and Future Directions
The study of CAA often relies on orthotopic mammary tumour models [1, 3]. In these models, TNBC cells and CAA are co-injected into the mammary fat pad of mice [1]. Observations from such experiments show that the presence of CAA accelerates primary tumour growth and promotes lung metastasis [1]. Histological analysis of the collected tissues confirms the activation of EMT and the remodelling of the immune environment [1, 3]. These organismal-level findings validate the observations made in cell culture systems [1].
The use of 6-well Transwell plates with polycarbonate membranes allows for the isolation of the effects of secreted factors from direct cell contact [1, 3]. This method has been essential for identifying the secretome of CAA and understanding its effect on cancer cell behavior [1, 3]. Furthermore, transcriptome sequencing of CAA versus normal adipocytes has revealed many differentially expressed genes that could serve as therapeutic targets [3, 7].
In summary, the interaction between TNBC cells and adipocytes is a major driver of tumour progression and immune escape [1, 2]. Adipocytes undergo a transformation into CAA, which then secrete cytokines like IL-6, IL-8, and G-CSF [2, 7]. These factors activate pathways like Stat3 and PI3K/AKT, leading to EMT and increased metastasis [1, 7]. CAA also reshape the TIME by upregulating PD-L1 and reducing T cell infiltration [1, 3]. Interventions such as EGCG or the targeting of exact chemokines like CXCL8 represent possible strategies to block these pro-tumour interactions [3, 4, 6]. The use of single-cell technologies continues to refine the knowledge of these cell populations and their parts in breast cancer biology [5].
References
Zhang, X., Zhong, S., Liu, S., Mu, X., Chen, G., & Chen, W. (2025). Cancer-associated adipocyte promote progression and immunosuppression in triple-negative breast cancer. PubMed Central. https://pmc.ncbi.nlm.nih.gov/articles/PMC13128427/
Yang, Z., Zeng, H., Li, J., et al. (2024). Dissecting the emerging role of cancer-associated adipocyte-derived cytokines in remodeling breast cancer progression. Heliyon, 10, e35200. https://doi.org/10.1016/j.heliyon.2024.e35200
Huang, R., Wang, Z., Hong, J., et al. (2023). Targeting cancer-associated adipocyte-derived CXCL8 inhibits triple-negative breast cancer progression and enhances the efficacy of anti-PD-1 immunotherapy. https://doi.org/10.21203/rs.3.rs-2419604/v1
Gonzalez Suarez, N., Fernandez-Marrero, Y., Hébert, M. P. A., et al. (2023). EGCG inhibits the inflammation and senescence inducing properties of MDA-MB-231 triple-negative breast cancer (TNBC) cells-derived extracellular vesicles in human adipose-derived mesenchymal stem cells. Cancer Cell International, 23. https://doi.org/10.1186/s12935-023-03087-2
Liu, S.-Q., Chen, D.-Y., Li, B., et al. (2023). Single-cell analysis of white adipose tissue reveals the tumor-promoting adipocyte subtypes. Journal of Translational Medicine, 21. https://doi.org/10.1186/s12967-023-04256-7
Gonzalez Suarez, N., Fernandez-Marrero, Y., Torabidastgerdooei, S., & Annabi, B. (2022). EGCG Prevents the Onset of an Inflammatory and Cancer-Associated Adipocyte-like Phenotype in Adipose-Derived Mesenchymal Stem/Stromal Cells in Response to the Triple-Negative Breast Cancer Secretome. Nutrients, 14, 1099. https://doi.org/10.3390/nu14051099
Liu, L., Wu, Y., Zhang, C., et al. (2020). Cancer-associated adipocyte-derived G-CSF promotes breast cancer malignancy via Stat3 signaling. Journal of Molecular Cell Biology, 12, 723-737. https://doi.org/10.1093/jmcb/mjaa016