Temperature control of microtubules assembly in S. pombe
Abstract
S. pombe is a very powerful organism to study microtubules assembly. The easy manipulation of fission yeast, the short dividing time and the existence of temperature-sensitive mutant strain allows researcher to dissect the role of microtubules in various aspect of S.pombe biology. Temperature is a controlling factor of microtubule assembly.
What if you could polymerize and depolymerize microtubules in a precise and ultra-fast fashion or combine microtubules studies with any of your temperature-sensitive mutant fission yeast? With our CherryTemp temperature controller you can switch from 5-45C in seconds, It has been featured in varios scientific publications, easy to use and fits easily with any microscope settings.
Microtubules and cellular processes
Microtubules are polymeric and very dynamic structures, which are part of the cytoskeleton of every cell. They play a crucial role in various cellular processes. During cell division they are important for chromosomes segregation in mitosis. They also ensure intra-cellular transport, nucleus and organelles movement. They participate in cell polarity and cellular migration. Microtubules, along with intermediate filament and actin filament can be seen as the bone and muscle of a cell, they really define cell architecture and shape. The role of microtubules in S.pombe cell division, growth and polarity has been very well documented (S.Martin, 2009; Piel and Tran, 2009).
Ultra fast temperature shift device for in vitro experiments under microscopy
Microtubules structure
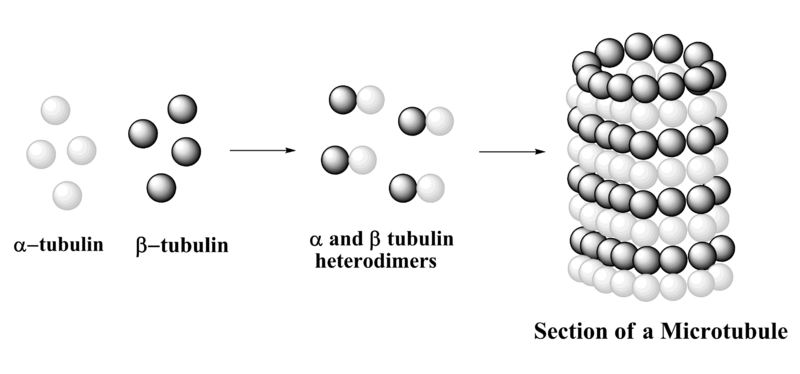
A microtubule is a hollow cylinder and a polarized non-covalent polymer, with a plus and a minus end. A microtubule is composed of 13 protofilaments, each of them composed of alfa and beta-tubulin heterodimers. There are three phases in microtubule assembly: a nucleation, an elongation and an equilibration phases. The nucleation phase consists in the formation of the alfa and beta-tubulin dimers. This phase is called nucleation as alfa and beta dimers are the “nucleus” chore components of microtubules. In S.pombe, microtubule nucleation occurs in perinuclear microtubules organizing center called spindle pole body. The elongation phase is a growing phase, during which tubulin dimers are added to the microtubule plus end allowing microtubules to elongate away from the MTOC. Finally when the concentration of polymerized actin is equivalent to that of cytoplasmic free actin, the microtubule has reached an equilibration phase.
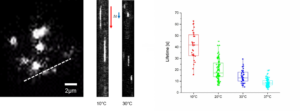
Temperature, microtubules and depolymerization
Microtubules are very dynamic structures, with a constant addition and removal of tubulin dimers at the microtubule plus end tip. When tubulin dimers depolymerize and microtubule start to shrink this is refer to as “catastrophe”. The switch from a catastrophe phase to a growing phase is called “rescue”phase.
A number of drugs can interfere with microtubule polymerization, among them colchicine has been widely used in laboratory. However, once microtubules are formed, their stability becomes temperature dependent (Lodish, 2000). Subjecting the cells to low temperature (6C) is enough to obtain microtubule depolymerization, along with relocation of microtubule-associated protein to the perinuclear domain. This cold-shock is reversible and with heat microtubules can polymerize again.
Interestingly, fishes swimming in cold water, posess tubulin which is cold resistant and can polymerize at very low temperature (-1.8C).
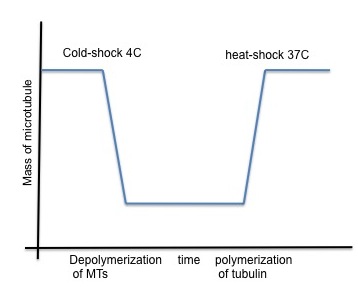
Microtubule and cell division:
During cell division, spindle microtubules are attached on one side to the spindle pole bodies and on the other side to chromosomes to pull them apart and into the future two daughter cells. During the cell cycle, if spindle microtubules are defective, the spindle checkpoint blocks mitosis progression. Taking advantage of a series of cold and heat-sensitive S.pombe beta-tubulin mutant along with temperature-shift experiments Paul Nurse’s team (Castagnetti et al., 2010) showed that while microtubules depolymerization triggers spindle checkpoint, it could quickly be bypassed with heat, and notably at high temperature. Indeed, after microtubules depolymerization, cytokinesis was delayed of 2 hours at 20 C, 45 min at 25C and no significant delays were observed at 32 C (Castagnetti et al., 2010). Following on these findings, they made the very interesting observation that without spindle microtubules, S.pombe can still undergo unusual nuclear fission.
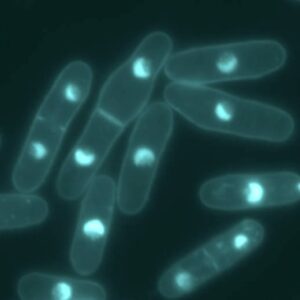
Studying microtubules with temperature-shift experiment
Microtubules are dynamic, they grow, shrink, undergo catastrophe in a very short time. Microtubule half-life is short, it ranges from 1 to 10 min. Doing cold shock to induce microtubules depolymerization followed by heat-shock to revert it and combining it with live cell imaging or video time-lapse can become very tricky. With CherryTemp you can do just that, switch temperature (5 to 45 C) in seconds, program cycle of polymerization and depolymerization. In a very nice study, Phong T Tran’s team (Fu et al., 2009) used CherryTemp to perform temperature-shift experiments and look at the specific role of microtubule associated protein and motor protein in spindle elongation. CherryTemp is a very easy and very efficient temperature controller device to perform ultra-fast activation, inactivation of temperature-sensitive proteins and dissect even further mechanisms that otherwise would be out of reach.
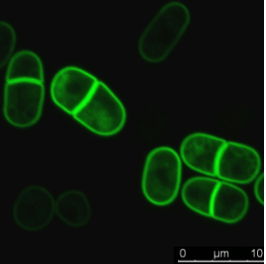
References
C.Fu, J J. Ward, I.Loiodice, G.Velve-Casquillas, FJ. Nedelec, and PT. Tran, Phospho-Regulated Interaction between Kinesin-6 Klp9p and Microtubule Bundler Ase1p Promotes Spindle Elongation, Developmental cell, 2009
S. Castagnetti, S. Oliferenko, P. Nurse Fission Yeast Cells Undergo Nuclear Division in the Absence of Spindle Microtubules, PLoS Biology, 2010
S. Martin Microtubule-dependent cell morphogenesis in the fission yeast, trends in Cell Biology, 2009
H.Lodish, Molecular Cell biology, WH Freeman edition.
M.Piel and PT. Tran Cell Shape and Cell Division in Fission Yeast , Current Biology 2009
FAQ
The organism S. pombe is applied in studies of microtubule assembly. Fission yeast is noted for its ease of manipulation. It also has a short dividing time. Additionally, temperature-sensitive mutant strains of this yeast exist. These characteristics permit researchers to examine the function of microtubules in different aspects of S. pombe biology. The assembly of microtubules is known to be a temperature-controlled factor. The function of microtubules in S. pombe cell division, growth, and polarity has been documented. The availability of temperature-sensitive mutants is particularly relevant for these types of investigations. This allows for experiments, such as temperature-shift experiments, to be conducted to understand cellular processes.
A microtubule is a hollow cylinder and a polarized non-covalent polymer. It has a plus and a minus end. The structure is composed of 13 protofilaments. Each of these protofilaments is composed of alpha and beta-tubulin heterodimers. There are three phases in microtubule assembly. The first is the nucleation phase, which involves the formation of the alpha and beta-tubulin dimers. In S. pombe, microtubule nucleation happens in perinuclear microtubules organizing centers, which are called spindle pole bodies. The second phase is elongation, a growing phase during which tubulin dimers are added to the microtubule plus end. This addition allows microtubules to elongate. Finally, an equilibration phase is reached when the concentration of polymerized material is equivalent to the free material in the cytoplasm.
Microtubules are structures that change, with a constant addition and removal of tubulin dimers at the microtubule plus end tip. When tubulin dimers depolymerize and microtubules begin to shrink, this is referred to as "catastrophe". The switch from a catastrophe phase to a growing phase is called a "rescue" phase. Once microtubules are formed, their stability becomes dependent on temperature. Subjecting the cells to a low temperature, such as 6°C, is sufficient to obtain microtubule depolymerization. This also causes the relocation of microtubule-associated proteins to the perinuclear domain. This cold-shock effect is reversible, and microtubules can polymerize again with heat. An exception is noted in fishes from cold water, which possess tubulin that is cold resistant and can polymerize at very low temperatures (-1.8°C).
Temperature-shift experiments have been used with temperature-sensitive S. pombe mutants. One study used a series of cold and heat-sensitive beta-tubulin mutants. It was shown that while microtubule depolymerization triggers the spindle checkpoint, this checkpoint could be quickly bypassed with heat. After microtubule depolymerization, cytokinesis was delayed for two hours at 20°C. The delay was only 45 minutes at 25°C, and no notable delays were observed at 32°C. Following these findings, an observation was made that S. pombe can still undergo "unusual nuclear fission" even in the absence of spindle microtubules. Other studies have also used temperature-shift experiments to examine the function of microtubule-associated proteins and motor proteins in spindle elongation.